
Page 107 - 2021_02-Haematologica-web
P. 107
MicroRNA-21 maintains HSC homeostasis
PDCD4 is involved in promoting NF-κB activation.44,45 However, in our study, we found that overexpression of PDCD4 in WT HSC by lentiviral transfection inhibited NF-κB activity, while knockdown of PDCD4 expression significantly improved the defects in miR-21-null HSC. These results elucidate that miR-21 sustains the NF-κB pathway, at least partly, by directly targeting PDCD4, which is consistent with the results of a previous study.33 However, whether miR-21 can regulate other target genes still needs further research. In addition, there is an oppos- ing point of view that miR-21 inhibits NF-κB activity,45 which may involve miR-21 regulation of physiological functions in a cellular context-dependent manner.
Ionizing radiation-caused hematopoietic cell death is primarily attributed to the generation of double-strand breaks, which are the most serious form of DNA damage.2 At present, the acute myelosuppression induced by irradi- ation can be temporarily treated using various hematopoi- etic growth factors that are able to accelerate the recovery of hematopoietic function by stimulating HSPC prolifera- tion and differentiation.46 However, an effective strategy to protect DNA from irradiation-induced damage in HSC is still lacking. Previous studies have revealed that miR-21 mediates resistance to radiation in many types of tumor cells, which is a great challenge for cancer radiotherapy.47 Furthermore, it has been shown that miR-21 can be stim-
ABC
DEF
GHI
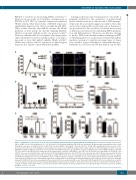
Figure 7. miR-21 protects hematopoietic stem cells from irradiation-induced damage by activating the NF-κB pathway. (A) Flow cytometric analysis of the percent- age of γ-H2AX+ cells at 0 (No irradiation) or 2, 6, 12, 24 h after 4.0 Gy total body irradiation (TBI) (n=6 mice per group). (B) Representative immunofluorescence plots showing the expression of γ-H2AX in Lin-Sca1+c-Kit+ (LSK) cells from miR-21fl/fl and miR-21∆/∆ mice 24 h after 4.0 Gy TBI. DAPI staining indicates the nucleus of cells. Scale bar represents 5 mm. (C) Quantitative real-time polymerase chain reaction (qRT-PCR) analysis of the relative expression of NF-κB target genes (Ier3, Xrcc5 and Gadd45b) in LSK from miR-21fl/fl and miR-21∆/∆ mice 5 h after 4.0 Gy TBI (n=3 mice per group). (D) The number of LSK in the bone marrow (BM) of miR-21fl/fl and miR- 21∆/∆ mice after 4.5 Gy TBI (n=6 mice per group). (E) The survival rates of miR-21fl/fl and miR-21∆/∆ mice after 6.5 Gy TBI (n=10 mice per group). (F, G) miR-21fl/fl and miR-21∆/∆ mice were treated with saline or thrombopoietin (TPO; 8 μg/kg) 30 min before 4.0 Gy TBI. Then, the expression of p-p65 in LSK was measured 40 min after irradiation (n=6 mice per group) (F), and the percentage of γ-H2AX+ LSK in these mice was measured by flow cytometry 24 h after irradiation (n=6 mice per group) (G). (H) Normal wild-type mice were treated with three doses of miR-21 agomir or negative control (NC) on consecutive days by tail intravenous injection. Twenty-four hours after the last injection, the relative miR-21 expression in LSK from these mice was detected by qRT-PCR (n=6 mice per group), and the expression of PDCD4 and p-p65 in LSK from these mice was analyzed by flow cytometry (n=6 mice per group). (I) Mice were treated with miR-21 agomir or NC as described before. Twenty- one hours after the last injection, mice were intraperitoneally injected with one dose of QNZ or control. Three hours later, mice were subjected to 4.0 Gy TBI. The per- centage of γ-H2AX+ cells in LSK from these mice was detected 24 h after irradiation by flow cytometry (n=6 mice per group). All data are shown as means ± standard deviation. *P<0.05, **P<0.01.
haematologica | 2021; 106(2)
421
