
Page 146 - 2021_02-Haematologica-web
P. 146
P.J. Hohensinner et al.
in such cells. In contrast, TF production in macrophages was not affected by inflammatory polarization with LPS and IFN-γ.
Differentiation of monocytes to macrophages can be induced by MCSF or GMCSF to simulate a chronic inflam- matory state or an acute event, respectively.24,25 Both cytokines were reported to result in differentiation of monocytes into mature macrophages, although GMCSF- derived macrophages display a higher proinflammatory potential.24 Here we show that human macrophages gener- ated from monocytes by stimulation with GMCSF pro- duce significantly more TF than macrophages generated from monocytes by stimulation with MCSF. We hypothe- size that the increased proinflammatory state induced by GMCSF might be responsible for this higher basal TF pro- tein level in GMCSF-derived macrophages as compared to their MCSF-derived counterparts. However, a significant increase in TF after alternative polarization was evident for macrophages differentiated from monocytes by either MCSF or GMCSF. Thus, this increase in TF was not dependent on the initial monocyte to macrophage differ- entiation by these colony-stimulating factors.
Besides coagulation, TF might also modulate the migra-
tory capacity of macrophages in a way similar to that observed for other cell types.22 Several integrins have already been reported to interact with TF.26 Interestingly, integrin α4 was associated with TF+ microvesicles shed from macrophages.27 This proinflammatory shedding was dependent on activation of caspase-1.28 Integrin α3 was associated with TF-dependent migration in keratinocytes.22 Our results indicate that integrin α3 is downregulated with polarization. We speculate that low levels of both integrin α3 and TF in proinflammatory macrophages might be responsible for the ability to form filopodia in laminin 5- positive areas.
Macrophages within inflammatory tissues or within tumors are derived from differentiated monocytes.29 Signals encountered in the microenvironment have the potential to specifically shape the developing macrophages.30 Previous results demonstrated that stimu- lating macrophages with pro- or anti-inflammatory cytokines leads to opposing transcriptional functional pro- grams and results in silencing or activation of genes causing a memory function of initial polarization conditions.31,32 Previously, we were able to demonstrate that an initial alternative activation of macrophages led to a subsequent
ABC
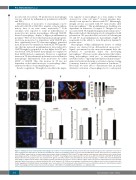
D
EF
Figure 6. Staining of tissue factor-positive macrophages in sections of colon carcinoma. (A) Tissue factor (TF) (red) and CD206 (green) were stained in colon cancer tissue using specific antibodies as described in the Methods. CD206+ macrophages positive for TF are indicated with white arrows. The boxed area is shown in more detail. (B) White arrows indicate acellular regions that showed double staining for CD206 and TF (orange), which could represent extracellular vesicles derived from macrophages positive for CD206 and TF. (C) Distribution of CD206+ macrophages was evaluated and scored in areas with low, medium, and high TF density. (D) Human atherosclerotic plaque tissue was stained for TF (green) and either CD206 (red) alternatively activated macrophages or CD80 (red) for proinflammatory macrophages. Adjusted TF intensity to macrophage intensity demonstrated an increase in TF in CD206+ regions. Values are given as adjusted tissue factor intensity (arbitrary units) mean values ± standard deviation (SD) (n=16 patients). (E, F) Mouse macrophages from atherosclerotic plaques were isolated as indicated in the Online Supplement. Proinflammatory macrophages were less positive for TF and showed reduced mean fluorescence intensity compared to alternatively activated CD206 macrophages (n=11). Values represent mean values ± SD. DAPI: 4′,6-diamidino-2-phenylindole; a.u.: arbitrary units.
460
haematologica | 2021; 106(2)
