
Page 28 - Haematologica3
P. 28
388
J.T. Butler et al.
and other cargo, and may explain the enhanced angio- genic and regenerative activity after hypoperfusion injury in distant tissues.53
Several groups have studied the release and function of EVs by BM stroma, such as endothelial cells and MSCs. Endothelial cells have been shown to generate EVs with pro-angiogenic effects through the actions of miR-126,54 and have been linked to age-related downregulation of osteogenic differentiation within the BM.55,56 More litera- ture exists on the release and function of EVs from BM- derived MSCs. Our group recently demonstrated the traf- ficking of EVs from BM-derived MSCs to hematopoietic cells influencing progenitor commitment.17 Other groups showed that MSC-derived EVs selectively promoted tumor growth in patients with multiple myeloma.21 Additionally MSC-derived EVs have been shown to regu- late angiogenic activity in endothelial cells, supporting the notion that BM MSC-derived EVs can regulate specific cell populations both within and outside of the hematopoietic compartment.57
In a recent study we showed that murine HSPCs (KSL: c-kit+/sca-1+/lineage-depleted) exposed to BM MSC- derived EVs in vitro prompted activation with myeloid pro- genitor biased expansion and a skewed hematopoietic repopulation potential.17 Remarkably, this process seemed to be dependent on Toll-like receptor signaling and could be specifically abrogated in HSPCs from TLR4 knockout
or MyD88 knockout animals (Figure 2A). EVs of all classes are also rich in lipid components, especially products of arachidonic acid metabolism, including prostaglandin E2.58 Considering the potent activity of prostaglandin E2 in reg- ulating HSC expansion and engraftment,59 it is tempting to speculate that EV-bound prostaglandin E2 released by MSCs contributes to this activity.60
The EV-mediated influence on hematopoiesis is not lim- ited to supportive stromal cells alone. Megakaryocytes have also been shown to impart regulatory control on HSPCs by releasing microvesicles to orchestrate specific cell-type commitment. Megakaryoctye-derived microvesicles are among the most abundant microvesicles in the circulation, and attach to HSPCs by interacting with ICAM-1, CD43, CD18 and CD11b epitopes. Upon cell surface contact, these microvesicles become internalized where megakaryocyte RNA appears to serve as the medi- ator of biological effects, as evidenced by a loss of function of megakaryocyte microvesicles following RNAase treat- ment. Functionally, the internalization of these megakaryocyte microvesicles was found to redirect the differentiation of HSPCs toward functional megakary- ocytes with limited effects on the phenotype of endothe- lial or stromal cells (Figure 2B).61
Several studies have demonstrated the importance of EV miRNA in regulating erythropoietic differentiation of HSPCs in both murine and human models.62 One recent
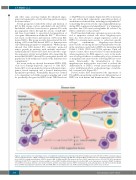
Figure 3. Current evidence for extracellular vesicle crosstalk in the leukemic microenvironment (A) EVs from AML blasts traffic miR-155 to HSPCs and down-regulate critical transcription factor, c-MYB, resulting in reduced differentiation potential. (B) AML EVs reprogram MSCs and stromal cells, and downregulate niche retention factor CXCL12 resulting in mobilization of HSPCs from the BM. (C) AML and MDS EVs promote the loss of HSPC supportive factors, CXCL12, SCF, IGF-1 through the trafficking of miR-7797 to supportive stroma, leading to reduced HSPC viability and hematopoietic potential. AML: acute myelogenous leukemia; ANGPT-1: angiopoi- etin 1; BM: bone marrow; CXCL12: C-X-C motif chemokine 12; EVs: extracellular vesicles; HSPCs: hematopoietic stem and progenitor cells; IGF-1: insulin-like growth factor 1; MDS: myelodysplastic syndrome; miR: microRNA; MSC: mesenchymal stem cell; PCBP1: poly(rc) binding protein 1; SCF: stem cell factor.
haematologica | 2018; 103(3)
