
Page 167 - 2019_06-Haematologica-web
P. 167
Low TLT-1 and collagen receptor α2 in FPD/AML
mature MK. Expression of mCherry fluorescent protein increased in both cases when TREML1_RR and ITGA2_RR were cloned upstream of the respective promoters and sig- nificantly decreased after specific deletion of RUNX1 binding sites (Figure 2E and F). These results clearly demonstrate that the identified sites are functional in mature MK and that TREML1 and ITGA2 are direct RUNX1 targets positively regulated by this transcription factor. mCherry reporter expression driven by TREML1_RR and ITGA2_RR harboring deleted RUNX1 sites did not reach the same level as for promoters alone, especially for ITGA2; this is probably because RUNX1 co- operates with other transcription factors. Indeed, ETS and
EVI1 binding sites and EVI1, SCL and ETS sites are identi- fied in TREML1_RR and ITGA2_RR regions, respectively. Moreover, at least one other site positive for H3K27Ac mark binds RUNX1 at position 52362118-52362307nt (Figure 2C). This site could also be involved in the regula- tion of ITGA2 by RUNX1.
TLT-1 is decreased in platelets and megakaryocytes in familial platelet disorder with predisposition to acute myelogenous leukemia
Considering that the main role of TLT-1 is related to platelet function, we next assessed the levels in platelets. First, we showed decreased TREML1 transcripts by qPCR
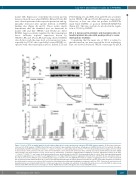
AB
C
D
Figure 3. Levels of TLT-1 in familial platelet disorder with predisposition to acute myelogenous leukemia (FPD/AML) patient samples. (A) Real-time polymerase chain reaction (PCR) analysis of TREML1 transcript levels normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) in platelets from patients of pedigree D harboring the T219Rfs*8 mutation (n=3): DII-1 ( !), DIII-1 ( !), DIII-3 ( !), and healthy subjects (n=10); **P<0.01. (B) (Left) Western blot analysis of TLT-1 content in platelet lysates from patients (n=3) and controls (n=4). Membranes were probed with anti-human TLT-1 antibody and reprobed with anti-beta (β)-actin. (Right) Optical density (OD) measurement of TLT-1/ b-actin ratio by densitometry. (C) Anti-TLT-1 antibody-induced inhibition of platelet aggregation. Patient (n=3): DII-1 ( !), DIII-1 ( !), DIII-3 ( !), and control (n=4) washed platelets were incubated with an anti-TLT-1 blocking antibody (Ab) or vehicle, then challenged with thrombin and platelet aggregation was recorded in a Lumi-Aggregometer. Percentage inhibition in platelet aggregation induced by the anti-TLT-1 Ab was calculated relative to the vehicle (set as 100%). Median values and interquartile range are shown; *P<0.05. (Right) Representative examples of aggregation traces of control and patient (DII-1) platelets incubated with anti-TLT-1 Ab or vehicle and challenged with thrombin. As patient platelets had reduced response to thrombin, thrombin concentration was titrated until a response was achieved. (D) Potentiation of thrombin-induced fibrinogen binding by recombinant soluble (rs) TLT-1. Patient (n=2) and control (n=2) washed platelets were incubated with rsTLT-1 or vehicle and exposed to increasing concentrations of thrombin. Mean fluorescence intensity (MFI) is expressed as arbitrary units (MFU). Median values and range are shown. (Right) Representative histograms and corresponding MFI values.
haematologica | 2019; 104(6)
1249
