
Page 29 - 2018_10-Haematologica-web
P. 29
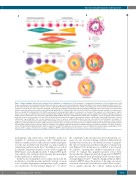
Normal and pathological erythropoiesis
A
B
C
D
Figure 1. Major pathways and molecules involved in the regulation of erythropoiesis. (A) Development of erythropoietic progenitor cells and erythroblasts. Early stages of erythropoietic development include primitive (p) and more mature (m) burst-forming unit erythroid cells (BFU-E) and colony-forming unit erythroid cells (CFU- E). The differentiation and maturation of these cells are regulated by broadly acting hematopoietic cytokines, including stem cell factor (SCF) and interleukin-3 (IL-3) and their receptors (R), the SCF receptor KIT and IL-3-R. Later stages are primarily regulated by erythropoietin (EPO) and EPO-R and are dependent on iron-metabolism and the interaction between the death receptor FAS and its ligand (FAS-L). Transferrin (Tf) and Tf-receptor-1 (TfR-1) are additional major regulators of erythropoiesis. Moreover, growth differentiating factor 11 (GDF11) and polymeric immunoglobulin A (IgA) are considered to be involved in the regulation of certain stages of erythro- poiesis. Later stages of erythropoiesis include proerythroblasts (ProEbl), basophilic (Baso) erythroblasts (type I and II), polychromatic (PolyC) erythroblasts and aci- dophilic (Acido) erythroblasts, also called late erythroblasts (Barbara Bain, personal communication to MCB). FAS-L and GDF11 are involved in the final maturation stage that leads to the generation of red cells (RC). (B) Erythroid blood island: macrophage surrounded by immature erythroblasts expressing FAS and more mature erythroblasts expressing FAS-L. The cell unit (island) acts in concert to promote erythroid differentiation and red cell production and maturation (rectangle). (C) Caspase activation during terminal erythroid differentiation: BID-dependent activation of caspase occurs in mitochondria. Various caspase targets are affected, includ- ing Rock-1, Lamin B and Acinus. However, GATA-1 is protected from caspase cleavage by heat shock protein 70 (HSP70). (D) Model of terminal erythroid differentiation and apoptosis regulated by the nuclear localization of HSP70. SCF and EPO trigger the proliferation and differentiation of erythroid progenitor cells (EPC). In RC pre- cursors (CFU-E through erythroblasts) EPO induces maturation as HSP70 translocates into the nucleus to protect GATA-1 from caspase-induced degradation. In the absence of EPO, caspase-3 induces the cleavage of GATA-1 as HSP70 cannot translocate to the nucleus, and, as a result, apoptosis occurs.
macrophages and enterocytes, and thereby make iron available to erythroid progenitor cells.33 Erythroferone, which is synthesized by early and late erythroid progeni- tor cells, has recently been described as a major regulator of hepcidin synthesis in the liver.34 In addition to its role in iron uptake, transferrin receptor-1 is also able to trigger the MAP kinase and phosphoinositide 3 kinase/AKT path- ways induced by erythropoietin and may thus participate in the regulation of erythropoiesis.35
In the model of regulation of red cell production by the modulation of erythropoietin sensitivity and rate of apop- tosis of erythroid progenitor and precursor cells, caspases are considered to be key enzymes (Figure 1).27-31 To add to
the complexity of the system, it has been shown that cas- pases are also activated during erythroid differentiation and that this activation is absolutely necessary to prepare the cells for enucleation and thus formation of mature red cells (Figure 1C,D). In this process, GATA-1 is not cleaved by caspases, ensuring that erythroid maturation is pre- served. To explain this apparent paradox it has been shown that the chaperone heat shock protein 70 (HSP70) plays an essential role in protecting GATA-1 from caspase cleavage (Figure 1C,D).25-31 Indeed, during erythroid differ- entiation HSP70 enters the nucleus at the time of caspase activation, and, at this level, interacts with GATA-1 to pro- tect it against cleavage. By contrast, during apoptosis
haematologica | 2018; 103(10)
1595
