
Page 43 - 2022_02-Haematologica-web
P. 43
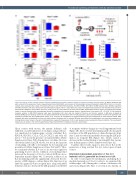
Functional switching of leukemic cells by stromal contact
AB
CD
Figure 3. Property of stem cell-like leukemic subsets exhibiting drug resistance without changes in leukemic initiating cell frequencies. (A) Murine leukemic cells were cultured in the presence of murine mesenchymal stromal cells (MSC), and numbers of surviving cells (annexinV- propidium iodide [PI]) in Sca-1(+) or Sca-1(-) subsets were measured after 2 days of treatment with Ara-C (100 nM for MN1 leukemia, 30 nM for H9M1, 500 nM for C1498 leukemic cells). (B) Mice engrafted with MN1 leukemic cells were treated with chemotherapeutic agents (Ara-C + doxorubicin) for 3 days by intraperitoneal injection at the indicated times and then examined for changes in the numbers of Sca-1(+) (LSK) or Sca-1(-) (LK) subsets in the bone marrow (BM). Experimental scheme (upper) and relative fold changes (lower) in the cell numbers after chemotherapy (chemoTx) compared to the control groups (mean ± standard error of the mean, n=5, *P<0.05). (C) Cell cycling of leukemic subsets in BM. MN1 leukemic cells were transplanted into mice and cell cycling of Sca-1(+) and Sca-1(-) subsets in BM were analyzed by Hoechst33342/pyronin staining. Shown are the representative flow cytometry plots with % of cell population (upper) and quantification of cells in G0 (quiescent cell population) and Non-G0 (G1/S/G2M) phase (lower) (n=3, *P<0.05). (D) Comparisons of leukemia-initiating cell (LIC) frequencies for each leukemic subset. MN1 leukemic cells were transplanted into mice and each subset of leukemic cells in recipient BM were sort-purified for transplantation into secondary recipients in a limiting dilution dose. Shown is the plot of limiting dilution analysis for frequencies of LIC in each leukemic subset analyzed by Poisson statistics. The resulting LIC frequencies are shown in the Online Supplementary Figure S2A with 95% Confidence Intervals in parenthesis.
Upon contact with stroma, the murine leukemic cells exhibited a notable induction of cytokines and growth fac- tors implicated in leukemogenic activity, including IL-4, PDGF-A, PDGF-D, CCL-2, CCL-5, CXCL-1 and stem cell factor,31-39 but not in the presence of transwell filters (Online Supplementary Figure S3B). Among those cytokines, IL-4 was selectively induced in LSK subsets, but not in the majority of remaining cells (LK) as determined by its transcript and protein level (Online Supplementary Figure S3C and D). Thus, we examined whether IL-4 acts as an autocrine signal for generating Sca-1(+) subsets. Addition of recombinant IL-4 increased the frequency of Sca-1(+) subsets (LSK) in a dose- dependent manner (Figure 4A). Conversely, addition of an IL-4-neutralizing antibody significantly decreased the fre- quency of LSK during co-culture (Figure 4A). Injection of an antibody against IL-4 into recipient mice along with MN1 leukemic cells also decreased the LSK population in the BM
of recipients without changes in overall engraftment levels (Figure 4B). Moreover, Il-4-neutralizing antibody abrogated resistance of the LSK population to chemotherapeutic drugs (Ara-C and doxorubicin), markedly decreasing the LSK pop- ulation in recipient BM (Figure 4C), which caused a decrease in the residual burden of surviving LIC that can ini- tiate leukemogenesis (Online Supplementary Figure S4).
Together, these results support a key role for IL-4 in the generation of drug resistant Sca-1(+) subset upon stromal contact.
Interleukin-4-dependent generation of Sca-1(+) leukemic cells is generated by stromal cross-talk
In order to investigate the mechanisms underlying IL-4- mediated generation of Sca-1(+) subsets, we examined the cellular target of IL-4 during the co-culture of leukemic cells and stroma (Figure 5A). First, to see if IL-4 acts directly on
haematologica | 2022; 107(2)
385
