
Page 262 - 2022_01-Haematologica-web
P. 262
A.O. Khan et al.
GH
I
J
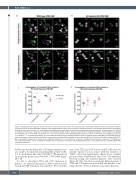
Figure 5. Polyglutamylation regulates spatial distribution of motor proteins in platelets and megakarycytes. (A and B) Quantification of the co-localization of dynein, as measured by the corrected Manders coefficient, with polyglutamylated tubulin over time shows a marked decrease in co-localization between these polymodified residues and the motor over time. (C) In fibrinogen and collagen spread cells, dynein is observed on the periphery of the spread cells, (D) while kinesin-1 is evident as a diffuse punctate stain. (E) Dynein co-localization with polyglutamylated residues decreases dramatically on platelet spreading in both fibrinogen and collagen (**P=0.0082 and ***P=0.0004 respectively). (F) A loss of co-localization with polyglutamylated tubulin is evident in fibrinogen and collagen spread cells (**P=0.0017, ****P<0.0001 respectively). (G) Immunofluorescence staining of induced pluripotent stem cell megacaryocytes (iPSC-MK) for polymodified tubulin, dynein, and kinesin-1 show the distribution of both motors along the length of the proplatelet shaft in wild-type (WT) cells. (H and I) In b1-tubulin knockout (KO) iPSC- MK, no proplatelet extensions are formed and a significant reduction in the co-localization of dynein to polyglutamlated residues is observed (*P=0.0166 and *P=0.0293 respectively), (J) with no significant change in the co-localization of kinesin-1 with polyglycylated tubulin. (n=3, standard deviation. Two-Way ANOVA with multiple comparisons.)
0.0004 respectively) (Figure 6D). CCP family enzymes were also found to be expressed in maturing MK, notably CCP1, 3, 4, 5, and 6 (Figure 6E), with CCP4 and CCP6 upregulated on proplatelet production (*P=0.0130, ***P=0.0009) (Figure 6F to H).
In order to investigate TTLL and CCP expression in platelets, we repeated this panel on RNA extracted from resting and CRP stimulated donor platelets. We found that
none of the TTLL and CCP observed in iPSC MK were con- sistently expressed across donors with the exception of TTLL7, a known polyglutamylase (Figure 6I, see complete gel in the Online Supplementary Figure S10).35 No differences between resting and activated platelets were observed (Figure 6I). This data shows a markedly different pattern of TTLL and CCP expression in both MK and platelets, corre- lating with the observed differences in polymodification.
254
haematologica | 2022; 107(1)
