
Page 126 - Haematologica Vol. 109 - July 2024
P. 126
ARTICLE - Cytotoxic reprogramming for BiTE immunotherapy M. Casey et al. AB
CD
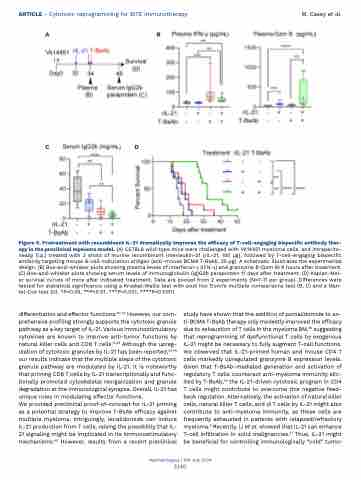
Figure 6. Pretreatment with recombinant IL-21 dramatically improves the efficacy of T-cell-engaging bispecific antibody ther- apy in the preclinical myeloma model. (A) C57BL6 wild-type mice were challenged with Vk14451 myeloma cells, and intraperito- neally (i.p.) treated with 2 shots of murine recombinant interleukin-21 (rIL-21, 100 μg), followed by T-cell-engaging bispecific antibody targeting mouse B-cell maturation antigen (anti-mouse BCMA T-BsAb, 25 μg). A schematic illustrates the experimental design. (B) Box-and-whisker plots showing plasma levels of interferon-γ (IFN-γ) and granzyme B (Gzm B) 6 hours after treatment. (C) Box-and-whisker plots showing serum levels of immunoglobulin (Ig)G2b paraprotein 11 days after treatment. (D) Kaplan-Mei- er survival curves of mice after indicated treatment. Data are pooled from 2 experiments (N=7-11 per group). Differences were tested for statistical significance using a Kruskal-Wallis test with post hoc Dunn’s multiple comparisons test (B, C) and a Man- tel-Cox test (D). *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
differentiation and effector functions.40-42 However, our com- prehensive profiling strongly supports the cytotoxic granule pathway as a key target of IL-21. Various immunostimulatory cytokines are known to improve anti-tumor functions by natural killer cells and CD8 T cells.14,43 Although the upreg- ulation of cytotoxic granules by IL-21 has been reported,42,44 our results indicate that the multiple steps of the cytotoxic granule pathway are modulated by IL-21. It is noteworthy that priming CD8 T cells by IL-21 transcriptionally and func- tionally promoted cytoskeletal reorganization and granule degradation at the immunological synapse. Overall, IL-21 has unique roles in modulating effector functions.
We provided preclinical proof-of-concept for IL-21 priming as a potential strategy to improve T-BsAb efficacy against multiple myeloma. Intriguingly, lenalidomide can induce IL-21 production from T cells, raising the possibility that IL- 21 signaling might be implicated in its immunostimulatory mechanisms.45 However, results from a recent preclinical
study have shown that the addition of pomalidomide to an- ti-BCMA T-BsAb therapy only modestly improved the efficacy due to exhaustion of T cells in the myeloma BM,46 suggesting that reprogramming of dysfunctional T cells by exogenous IL-21 might be necessary to fully augment T-cell functions. We observed that IL-21-primed human and mouse CD4 T cells markedly upregulated granzyme B expression levels. Given that T-BsAb-mediated generation and activation of regulatory T cells counteract anti-myeloma immunity elic- ited by T-BsAb,24 the IL-21-driven cytotoxic program in CD4 T cells might contribute to overcome this negative feed- back regulation. Alternatively, the activation of natural killer cells, natural killer T cells, and γδ T cells by IL-21 might also contribute to anti-myeloma immunity, as these cells are frequently exhausted in patients with relapsed/refractory myeloma.9 Recently, Li et al. showed that IL-21 can enhance T-cell infiltration in solid malignancies.47 Thus, IL-21 might be beneficial for controlling immunologically “cold” tumor
Haematologica | 109 July 2024
2140
